
Page 33 - Academic Press Encyclopedia of Physical Science and Technology 3rd Molecular Biology
P. 33
P1: GNH Revised Pages
Encyclopedia of Physical Science and Technology EN002G-104 May 17, 2001 20:53
Chromatin Structure and Modification 825
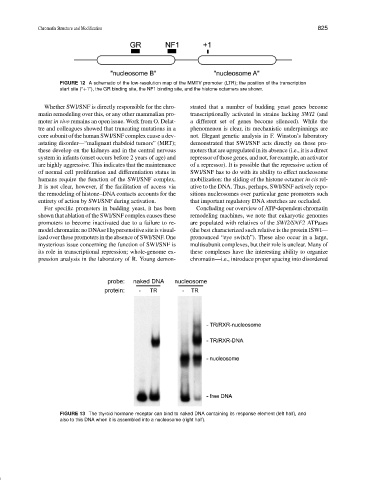
FIGURE 12 A schematic of the low-resolution map of the MMTV promoter (LTR); the position of the transcription
start site (“+1”), the GR binding site, the NF1 binding site, and the histone octamers are shown.
Whether SWI/SNF is directly responsible for the chro- strated that a number of budding yeast genes become
matin remodeling over this, or any other mammalian pro- transcriptionally activated in strains lacking SWI2 (and
moter in vivo remains an open issue. Work from O. Delat- a different set of genes become silenced). While the
tre and colleagues showed that truncating mutations in a phenomenon is clear, its mechanistic underpinnings are
core subunit of the human SWI/SNF complex cause a dev- not. Elegant genetic analysis in F. Winston’s laboratory
astating disorder—“malignant rhabdoid tumors” (MRT); demonstrated that SWI/SNF acts directly on those pro-
these develop on the kidneys and in the central nervous moters that are upregulated in its absence (i.e., it is a direct
system in infants (onset occurs before 2 years of age) and repressor of those genes, and not, for example, an activator
are highly aggressive. This indicates that the maintenance of a repressor). It is possible that the repressive action of
of normal cell proliferation and differentiation status in SWI/SNF has to do with its ability to effect nucleosome
humans require the function of the SWI/SNF complex. mobilization: the sliding of the histone octamer in cis rel-
It is not clear, however, if the facilitation of access via ative to the DNA. Thus, perhaps, SWI/SNF actively repo-
the remodeling of histone–DNA contacts accounts for the sitions nucleosomes over particular gene promoters such
entirety of action by SWI/SNF during activation. that important regulatory DNA stretches are occluded.
For specific promoters in budding yeast, it has been Concluding our overview of ATP-dependent chromatin
shown that ablation of the SWI/SNF complex causes these remodeling machines, we note that eukaryotic genomes
promoters to become inactivated due to a failure to re- are populated with relatives of the SWI2/SNF2 ATPases
modelchromatin:noDNAseIhypersensitivesiteisvisual- (the best characterized such relative is the protein ISWI—
ized over these promoters in the absence of SWI/SNF. One pronounced “eye switch”). These also occur in a large,
mysterious issue concerning the function of SWI/SNF is multisubunit complexes, but their role is unclear. Many of
its role in transcriptional repression; whole-genome ex- these complexes have the interesting ability to organize
pression analysis in the laboratory of R. Young demon- chromatin—i.e., introduce proper spacing into disordered
FIGURE 13 The thyroid hormone receptor can bind to naked DNA containing its response element (left half), and
also to this DNA when it is assembled into a nucleosome (right half).