
Page 379 - Introduction to Paleobiology and The Fossil Record
P. 379
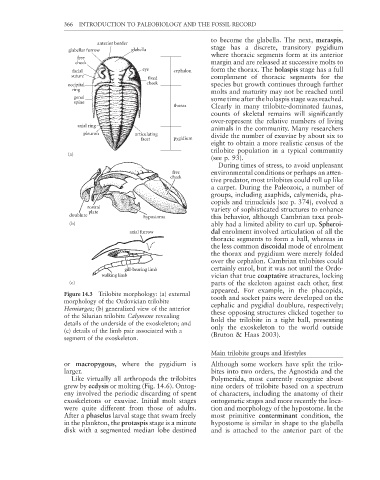
366 INTRODUCTION TO PALEOBIOLOGY AND THE FOSSIL RECORD
to become the glabella. The next, meraspis,
anterior border
glabellar furrow glabella stage has a discrete, transitory pygidium
where thoracic segments form at its anterior
free
cheek margin and are released at successive molts to
facial eye cephalon form the thorax. The holaspis stage has a full
suture complement of thoracic segments for the
fixed
occipital cheek species but growth continues through further
ring
molts and maturity may not be reached until
genal some time after the holaspis stage was reached.
spine
thorax Clearly in many trilobite-dominated faunas,
counts of skeletal remains will signifi cantly
over-represent the relative numbers of living
axial ring
animals in the community. Many researchers
pleuron articulating divide the number of exuviae by about six to
facet pygidium
eight to obtain a more realistic census of the
trilobite population in a typical community
(a)
(see p. 93).
During times of stress, to avoid unpleasant
free environmental conditions or perhaps an atten-
cheek
tive predator, most trilobites could roll up like
a carpet. During the Paleozoic, a number of
groups, including asaphids, calymenids, pha-
copids and trinucleids (see p. 374), evolved a
rostral variety of sophisticated structures to enhance
plate
doublure hypostoma this behavior, although Cambrian taxa prob-
(b) ably had a limited ability to curl up. Spheroi-
axial furrow dal enrolment involved articulation of all the
thoracic segments to form a ball, whereas in
the less common discoidal mode of enrolment
the thorax and pygidium were merely folded
over the cephalon. Cambrian trilobites could
gill-bearing limb certainly enrol, but it was not until the Ordo-
walking limb vician that true coaptative structures, locking
(c) parts of the skeleton against each other, fi rst
appeared. For example, in the phacopids,
Figure 14.3 Trilobite morphology: (a) external tooth and socket pairs were developed on the
morphology of the Ordovician trilobite cephalic and pygidial doublure, respectively;
Hemiarges; (b) generalized view of the anterior these opposing structures clicked together to
of the Silurian trilobite Calymene revealing hold the trilobite in a tight ball, presenting
details of the underside of the exoskeleton; and only the exoskeleton to the world outside
(c) details of the limb pair associated with a (Bruton & Haas 2003).
segment of the exoskeleton.
Main trilobite groups and lifestyles
or macropygous, where the pygidium is Although some workers have split the trilo-
larger. bites into two orders, the Agnostida and the
Like virtually all arthropods the trilobites Polymerida, most currently recognize about
grew by ecdysis or molting (Fig. 14.6). Ontog- nine orders of trilobite based on a spectrum
eny involved the periodic discarding of spent of characters, including the anatomy of their
exoskeletons or exuviae. Initial molt stages ontogenetic stages and more recently the loca-
were quite different from those of adults. tion and morphology of the hypostome. In the
After a phaselus larval stage that swam freely most primitive conterminant condition, the
in the plankton, the protaspis stage is a minute hypostome is similar in shape to the glabella
disk with a segmented median lobe destined and is attached to the anterior part of the