
Page 177 - Academic Press Encyclopedia of Physical Science and Technology 3rd BioTechnology
P. 177
P1: GKY/GLQ/GTK P2: GLM Final Pages
Encyclopedia of Physical Science and Technology en009I-422 July 6, 2001 19:57
Metabolic Engineering 405
consumption, e.g., fungal cells that do not produce afla- is therefore necessary to evaluate which of the two vari-
toxins are used for production of food grade enzymes, and ables is most important. Similarly, an improvement in the
E. coli strains that do not produce endotoxins are used for productivity through metabolic engineering does not nec-
production of pharmaceuticals. The second reason may essarily lead to an improvement in the yield of product on
especially be problematic if the by-product has properties the substrate. The simple model pathways shown in Fig. 8
similar to those of the desired product, since separation illustrate this. Here the substrate can be converted both to
of the active product then requires very efficient separa- the product of interest and to a by-product—in practice
tion principles, which are often costly. In some cases the there is often several different by-products. In this path-
by-product may also cause other types of problems, e.g., way the yield is given as the ratio between the fluxes J 2
inactivation of the desired product. Loss of carbon in the and J 1 . Clearly the yield is not changed if the flux distri-
by-product is mainly a problem in connection with the pro- bution around the branch point metabolite I is constant,
duction of low value added products, and here the strate- i.e., independently of the size of flux J 1 the ratios J 2 /J 1
gies discussed in the following section may be applied. and J 3 /J 1 are constant, whereas the productivity clearly
increases when J 1 increases. The flux distribution around
the branch point metabolite I depend on the flexibility or
G. Improvement of Yield or Productivity
rigidity of the network. The flexibility of a branch point
In the production of low value added products like ethanol, is determined by the affinities of the enzymes competing
lactic acid, citric acid, amino acids, and many antibiotics, for this metabolite and the concentration of the metabo-
the yield of product on the substrate and the productivity lite. If the metabolite concentration is much below the K m
are the most important design variables to optimize. Yield values of the two enzymes, then the flux distribution is
impacts primarily the cost of raw materials and is affected likely to be maintained if the supply of the metabolite via
by the metabolic fluxes, i.e., an increase in the yield is ob- flux J 1 increases. However, if both enzymes are saturated
tained through redirection of the fluxes toward the desired with the metabolite, then an increase in the flux J 1 will
product. Productivity, on the other hand, is of importance have no effect on either the yield or the productivity. In
when the capital costs are of importance, and it can be this case one may increase the yield and the productivity
improved by amplification of the metabolic fluxes. In or- through increasing the flux J 2 , e.g., by overexpressing the
der to obtain a high yield and productivity it is necessary enzyme E 2 . However, if an increase in E j results in a de-
to direct the carbon fluxes from the substrate toward the crease in the concentration of I below the K m values of
metabolite of interest at a high rate. This often requires en- the enzymes E j and E k , the consequence may be an al-
gineering of the central carbon metabolism, which is dif- tered flux distribution. Thus, even in this simple pathway
ficult due to the tight regulation in this part of the cellular structure it is difficult to design the appropriate strategy
metabolism. for increasing the yield or productivity without knowl-
For most processes it is not possible to operate the pro- edge of the kinetics of the enzymes and the metabolite
cess such that both the yield of product on the substrate concentration. The complexity increases further if there
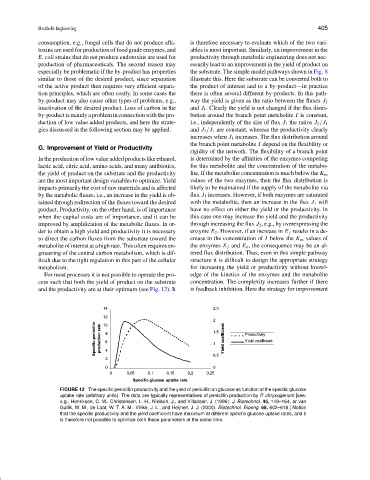
and the productivity are at their optimum (see Fig. 12). It is feedback inhibition. Here the strategy for improvement
FIGURE 12 The specific penicillin productivity and the yield of penicillin on glucose as function of the specific glucose
uptake rate (arbitrary units). The data are typically representatives of penicillin production by P. chrysogenum [see,
e.g., Henriksen, C. M., Christensen, L. H., Nielsen, J., and Villadsen, J. (1996). J. Biotechnol. 45, 149–164, or van
Gullik, W. M., de Laat, W. T. A. M., Vinke, J. L., and Heijnen, J. J. (2000). Biotechnol. Bioeng. 68, 602–618.] Notice
that the specific productivity and the yield coefficient have maximum at different specific glucose uptake rates, and it
is therefore not possible to optimize both these parameters at the same time.