
Page 112 - The Biochemistry of Inorganic Polyphosphates
P. 112
WU095/Kulaev
WU095-07
Functions of polyphosphate and polyphosphate-dependent enzymes
96 March 9, 2004 15:39 Char Count= 0
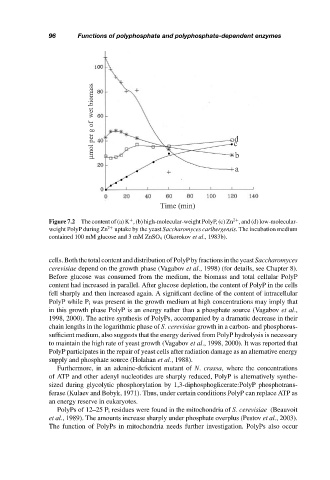
µmol per g of wet biomass c
d
b
a
Time (min)
Figure 7.2 The content of (a) K , (b) high-molecular-weight PolyP, (c) Zn , and (d) low-molecular-
+
2+
weight PolyP during Zn 2+ uptake by the yeast Saccharomyces carlbergensis. The incubation medium
contained 100 mM glucose and 3 mM ZnSO 4 (Okorokov et al., 1983b).
cells.BoththetotalcontentanddistributionofPolyPbyfractionsintheyeastSaccharomyces
cerevisiae depend on the growth phase (Vagabov et al., 1998) (for details, see Chapter 8).
Before glucose was consumed from the medium, the biomass and total cellular PolyP
content had increased in parallel. After glucose depletion, the content of PolyP in the cells
fell sharply and then increased again. A significant decline of the content of intracellular
PolyP while P i was present in the growth medium at high concentrations may imply that
in this growth phase PolyP is an energy rather than a phosphate source (Vagabov et al.,
1998, 2000). The active synthesis of PolyPs, accompanied by a dramatic decrease in their
chain lengths in the logarithmic phase of S. cerevisiae growth in a carbon- and phosphorus-
sufficient medium, also suggests that the energy derived from PolyP hydrolysis is necessary
to maintain the high rate of yeast growth (Vagabov et al., 1998, 2000). It was reported that
PolyP participates in the repair of yeast cells after radiation damage as an alternative energy
supply and phosphate source (Holahan et al., 1988).
Furthermore, in an adenine-deficient mutant of N. crassa, where the concentrations
of ATP and other adenyl nucleotides are sharply reduced, PolyP is alternatively synthe-
sized during glycolytic phosphorylation by 1,3-diphosphoglicerate:PolyP phosphotrans-
ferase (Kulaev and Bobyk, 1971). Thus, under certain conditions PolyP can replace ATP as
an energy reserve in eukaryotes.
PolyPs of 12–25 P i residues were found in the mitochondria of S. cerevisiae (Beauvoit
et al., 1989). The amounts increase sharply under phosphate overplus (Pestov et al., 2003).
The function of PolyPs in mitochondria needs further investigation. PolyPs also occur