
Page 127 - The Biochemistry of Inorganic Polyphosphates
P. 127
March 9, 2004
Char Count= 0
15:39
WU095/Kulaev
WU095-07
Gene activity control, development and stress response 111
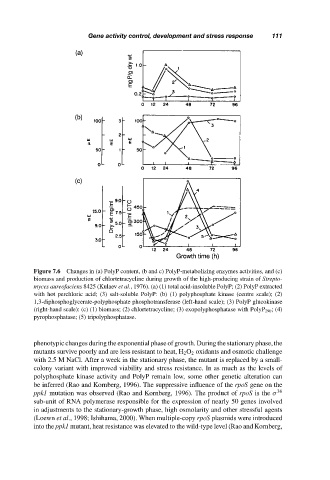
Figure 7.6 Changes in (a) PolyP content, (b and c) PolyP-metabolizing enzymes activities, and (c)
biomass and production of chlortetracycline during growth of the high-producing strain of Strepto-
myces aureofaciens 8425 (Kulaev et al., 1976). (a) (1) total acid-insoluble PolyP; (2) PolyP extracted
with hot perchloric acid; (3) salt-soluble PolyP: (b) (1) polyphosphate kinase (centre scale); (2)
1,3-diphosphoglycerate-polyphosphate phosphotransferase (left-hand scale); (3) PolyP glucokinase
(right-hand scale): (c) (1) biomass; (2) chlortetracycline; (3) exopolyphosphatase with PolyP 290 ; (4)
pyrophosphatase; (5) tripolyphosphatase.
phenotypic changes during the exponential phase of growth. During the stationary phase, the
mutants survive poorly and are less resistant to heat, H 2 O 2 oxidants and osmotic challenge
with 2.5 M NaCl. After a week in the stationary phase, the mutant is replaced by a small-
colony variant with improved viability and stress resistance. In as much as the levels of
polyphosphate kinase activity and PolyP remain low, some other genetic alteration can
be inferred (Rao and Kornberg, 1996). The suppressive influence of the rpoS gene on the
ppk1 mutation was observed (Rao and Kornberg, 1996). The product of rpoS is the σ 38
sub-unit of RNA polymerase responsible for the expression of nearly 50 genes involved
in adjustments to the stationary-growth phase, high osmolarity and other stressful agents
(Loewn et al., 1998; Ishihama, 2000). When multiple-copy rpoS plasmids were introduced
into the ppk1 mutant, heat resistance was elevated to the wild-type level (Rao and Kornberg,