
Page 60 - Biomimetics : Biologically Inspired Technologies
P. 60
Bar-Cohen : Biomimetics: Biologically Inspired Technologies DK3163_c002 Final Proof page 46 21.9.2005 11:39am
46 Biomimetics: Biologically Inspired Technologies
Furthermore, there are several structural proteins present in the M-line (i.e., M-protein, myomesin)
and the A-band (C-protein) that presumably keep the myosin filaments in register during contraction.
Sarcomere force production stems from the interaction between the actin and myosin filaments.
In vertebrate sarcomeres, six actin filaments surround each myosin filament. Myosin filaments of
vertebrates consist of approximately 100 myosin molecules, each shaped like a golf club with a
double head. The myosin heads protrude from the core of the filament towards the surrounding actin
filaments. Actin filaments consist of two helical strands of F-actin twined together like a bead
necklace. On each of the beads is a site where myosin can bind. Binding is regulated by the
2þ
configuration of the proteins troponin and tropomyosin, which is controlled by Ca . When a
myosin head attaches to an actin-binding site, it undergoes a conformational change resulting in the
development of force and sliding of the actin and myosin filaments along each other. Under the
influence of adenosine triphosphate (ATP), the crossbridge detaches again. Pumping back calcium
ions into the SR via ATP-consuming calcium pumps triggers the relaxation. The formation of
connections between myosin and actin is a stochastic process and it is known as the crossbridge
theory (Huxley, 1957, 2000).
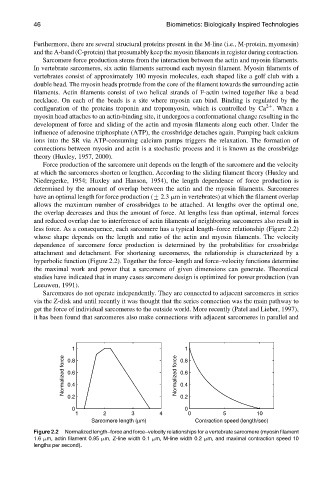
Force production of the sarcomere unit depends on the length of the sarcomere and the velocity
at which the sarcomeres shorten or lengthen. According to the sliding filament theory (Huxley and
Niedergerke, 1954; Huxley and Hanson, 1954), the length dependence of force production is
determined by the amount of overlap between the actin and the myosin filaments. Sarcomeres
have an optimal length for force production (+ 2.3 mm in vertebrates) at which the filament overlap
allows the maximum number of crossbridges to be attached. At lengths over the optimal one,
the overlap decreases and thus the amount of force. At lengths less than optimal, internal forces
and reduced overlap due to interference of actin filaments of neighboring sarcomeres also result in
less force. As a consequence, each sarcomere has a typical length–force relationship (Figure 2.2)
whose shape depends on the length and ratio of the actin and myosin filaments. The velocity
dependence of sarcomere force production is determined by the probabilities for crossbridge
attachment and detachment. For shortening sarcomeres, the relationship is characterized by a
hyperbolic function (Figure 2.2). Together the force–length and force–velocity functions determine
the maximal work and power that a sarcomere of given dimensions can generate. Theoretical
studies have indicated that in many cases sarcomere design is optimized for power production (van
Leeuwen, 1991).
Sarcomeres do not operate independently. They are connected to adjacent sarcomeres in series
via the Z-disk and until recently it was thought that the series connection was the main pathway to
get the force of individual sarcomeres to the outside world. More recently (Patel and Lieber, 1997),
it has been found that sarcomeres also make connections with adjacent sarcomeres in parallel and
1
1 0.8
Normalized force 0.6 Normalized force 0.6
0.8
0.4
0.4
0.2 0.2
0 0
1 2 3 4 0 5 10
Sarcomere length (µm) Contraction speed (length/sec)
Figure 2.2 Normalized length–force and force–velocity relationships for a vertebrate sarcomere (myosin filament
1.6 mm, actin filament 0.95 mm, Z-line width 0.1 mm, M-line width 0.2 mm, and maximal contraction speed 10
lengths per second).