
Page 87 - Introduction to Paleobiology and The Fossil Record
P. 87
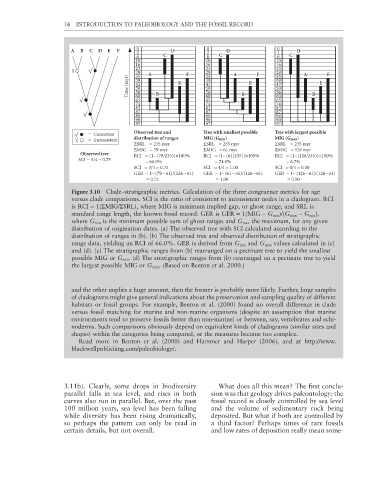
74 INTRODUCTION TO PALEOBIOLOGY AND THE FOSSIL RECORD
A B C D E F 0 D 0 D 0 D
2 2 2
6 C 6 C 6 C
10 10 10
16 16 16
23 23 23
29 A F 29 A F 29 A F
35
35
35
Time (myr) 39 E 39 E 39 E
42
42
42
50
50
50
56
56
56
60
65
65
65 B 60 B 60 B
74 74 74
83 83 83
87 87 87
88 88 88
90 90 90
97 97 97
= Consistent Observed tree and Tree with smallest possible Tree with largest possible
distribution of ranges MIG (G min) MIG (G max)
= Inconsistent
ΣSRL = 235 myr ΣSRL = 235 myr ΣSRL = 235 myr
ΣMIG = 79 myr ΣMIG = 61 myr ΣMIG = 126 myr
Observed tree
RCI = (1– (79/235))×100% RCI = (1– (61/235))×100% RCI = (1– (126/235))×100%
SCI = 3/4 = 0.75 = 66.0% = 74.0% = 6.7%
SCI = 3/4 = 0.75 SCI = 4/4 = 1.00 SCI = 0/4 = 0.00
GER = 1– (79 – 61)/(126 – 61) GER = 1– (61 – 61)/(126 – 61) GER = 1– (126 – 61)/(126 – 61)
= 0.72 = 1.00 = 0.00
Figure 3.10 Clade–stratigraphic metrics. Calculation of the three congruence metrics for age
versus clade comparisons. SCI is the ratio of consistent to inconsistent nodes in a cladogram. RCI
is RCI = 1(ΣMIG/ΣSRL), where MIG is minimum implied gap, or ghost range, and SRL is
standard range length, the known fossil record. GER is GER = 1(MIG − G min )/(G max − G min ),
where G min is the minimum possible sum of ghost ranges and G max the maximum, for any given
distribution of origination dates. (a) The observed tree with SCI calculated according to the
distribution of ranges in (b). (b) The observed tree and observed distribution of stratigraphic
range data, yielding an RCI of 66.0%. GER is derived from G min and G max values calculated in (c)
and (d). (c) The stratigraphic ranges from (b) rearranged on a pectinate tree to yield the smallest
possible MIG or G min . (d) The stratigraphic ranges from (b) rearranged on a pectinate tree to yield
the largest possible MIG or G max . (Based on Benton et al. 2000.)
and the other implies a huge amount, then the former is probably more likely. Further, large samples
of cladograms might give general indications about the preservation and sampling quality of different
habitats or fossil groups. For example, Benton et al. (2000) found no overall difference in clade
versus fossil matching for marine and non-marine organisms (despite an assumption that marine
environments tend to preserve fossils better than non-marine) or between, say, vertebrates and echi-
noderms. Such comparisons obviously depend on equivalent kinds of cladograms (similar sizes and
shapes) within the categories being compared, or the measures become too complex.
Read more in Benton et al. (2000) and Hammer and Harper (2006), and at http://www.
blackwellpublishing.com/paleobiology/.
3.11b). Clearly, some drops in biodiversity What does all this mean? The fi rst conclu-
parallel falls in sea level, and rises in both sion was that geology drives paleontology: the
curves also run in parallel. But, over the past fossil record is closely controlled by sea level
100 million years, sea level has been falling and the volume of sedimentary rock being
while diversity has been rising dramatically, deposited. But what if both are controlled by
so perhaps the pattern can only be read in a third factor? Perhaps times of rare fossils
certain details, but not overall. and low rates of deposition really mean some-