
Page 180 - The Biochemistry of Inorganic Polyphosphates
P. 180
WU095/Kulaev
WU095-08
Peculiarities of polyphosphate metabolism
164 March 9, 2004 20:32 Char Count= 0
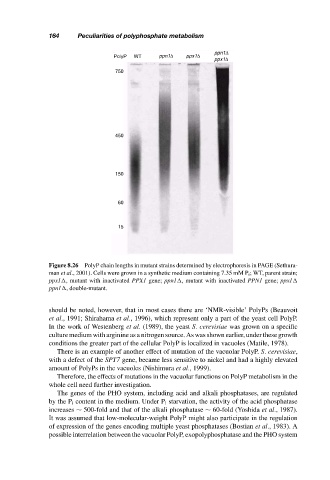
ppn1∆
PolyP WT ppn1∆ ppx1∆
ppx1∆
750
450
150
60
15
Figure 8.26 PolyP chain lengths in mutant strains determined by electrophoresis in PAGE (Sethura-
man et al., 2001). Cells were grown in a synthetic medium containing 7.35 mM P i ; WT, parent strain;
ppx1 , mutant with inactivated PPX1 gene; ppn1 , mutant with inactivated PPN1 gene; ppx1
ppn1 , double-mutant.
should be noted, however, that in most cases there are ‘NMR-visible’ PolyPs (Beauvoit
et al., 1991; Shirahama et al., 1996), which represent only a part of the yeast cell PolyP.
In the work of Westenberg et al. (1989), the yeast S. cerevisiae was grown on a specific
culture medium with arginine as a nitrogen source. As was shown earlier, under these growth
conditions the greater part of the cellular PolyP is localized in vacuoles (Matile, 1978).
There is an example of another effect of mutation of the vacuolar PolyP. S. cerevisiae,
with a defect of the SPT7 gene, became less sensitive to nickel and had a highly elevated
amount of PolyPs in the vacuoles (Nishimura et al., 1999).
Therefore, the effects of mutations in the vacuolar functions on PolyP metabolism in the
whole cell need further investigation.
The genes of the PHO system, including acid and alkali phosphatases, are regulated
by the P i content in the medium. Under P i starvation, the activity of the acid phosphatase
increases ∼ 500-fold and that of the alkali phosphatase ∼ 60-fold (Yoshida et al., 1987).
It was assumed that low-molecular-weight PolyP might also participate in the regulation
of expression of the genes encoding multiple yeast phosphatases (Bostian et al., 1983). A
possible interrelation between the vacuolar PolyP, exopolyphosphatase and the PHO system