
Page 18 - Carbonate Sedimentology and Sequence Stratigraphy
P. 18
CHAPTER 1: ESSENTIALS OF NEIGHBORING DISCIPLINES 9
dN = rN grow more and reproduce less or vice versa (Townsend et
dt al., 2003). In a very influential book, MacArthur and Wil-
son (1967) introduced two extreme life strategies that corre-
spond to these alternatives. There are organisms, such as
dN = rN (1- ) weeds, that quickly appear in newly opened habitats, mul-
N
K dt K tiply rapidly, are short-lived and relatively small. They are
called r-strategists because they spend most of their life in
the r-dominated, nearly exponential part of the population
growth curve in Fig. 1.13. At the other end of the spectrum
N
are organisms, such as large trees in a forest, that live in
environments of intense competition for limited resources.
These organisms grow fast, live long, and reproduce slowly
and relatively late in their life; they are called K-strategists
as most of their life is spent in the K-dominated part of the
0
Time (t) growth curve.
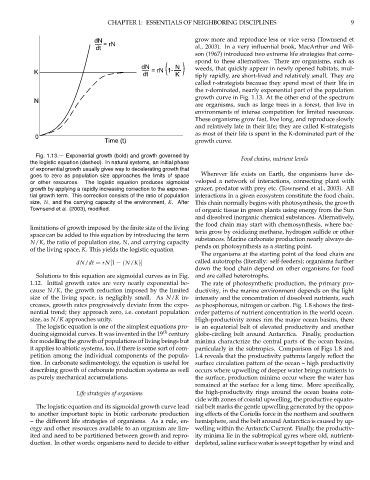
Fig. 1.13.— Exponential growth (bold) and growth governed by Food chains, nutrient levels
the logistic equation (dashed). In natural systems, an initial phase
of exponential growth usually gives way to decelerating growth that
goes to zero as population size approaches the limits of space Wherever life exists on Earth, the organisms have de-
or other resources. The logistic equation produces sigmoidal veloped a network of interactions, connecting plant with
growth by applying a rapidly increasing correction to the exponen- grazer, predator with prey etc. (Townsend et al., 2003). All
tial growth term. This correction consists of the ratio of population interactions in a given ecosystem constitute the food chain.
size, N, and the carrying capacity of the environment, K. After This chain normally begins with photosynthesis, the growth
Townsend et al. (2003), modified. of organic tissue in green plants using energy from the Sun
and dissolved inorganic chemical substances. Alternatively,
the food chain may start with chemosynthesis, where bac-
limitations of growth imposed by the finite size of the living
space can be added to this equation by introducing the term teria grow by oxidizing methane, hydrogen sulfide or other
N/K, the ratio of population size, N, and carrying capacity substances. Marine carbonate production nearly always de-
of the living space, K. This yields the logistic equation pends on photosynthesis as a starting point.
The organisms at the starting point of the food chain are
dN/dt = rN[1 − (N/K)] called autotrophs (literally: self-feeders); organisms further
down the food chain depend on other organisms for food
Solutions to this equation are sigmoidal curves as in Fig. and are called heterotrophs.
1.12. Initial growth rates are very nearly exponential be- The rate of photosynthetic production, the primary pro-
cause N/K, the growth reduction imposed by the limited ductivity, in the marine environment depends on the light
size of the living space, is negligibly small. As N/K in- intensity and the concentration of dissolved nutrients, such
creases, growth rates progressively deviate from the expo- as phosphorous, nitrogen or carbon. Fig. 1.8 shows the first-
nential trend; they approach zero, i.e. constant population order patterns of nutrient concentration in the world ocean.
size, as N/K approaches unity. High-productivity zones rim the major ocean basins, there
The logistic equation is one of the simplest equations pro- is an equatorial belt of elevated productivity and another
th
ducing sigmoidal curves. It was invented in the 19 century globe-circling belt around Antarctica. Finally, production
for modelling the growth of populations of living beings but minima characterize the central parts of the ocean basins,
it applies to abiotic systems, too, if there is some sort of com- particularly in the subtropics. Comparison of Figs 1.8 and
petition among the individual components of the popula- 1.4 reveals that the productivity patterns largely reflect the
tion. In carbonate sedimentology, the equation is useful for surface circulation pattern of the ocean – high productivity
describing growth of carbonate production systems as well occurs where upwelling of deeper water brings nutrients to
as purely mechanical accumulations. the surface, production minima occur where the water has
remained at the surface for a long time. More specifically,
Life strategies of organisms the high-productivity rings around the ocean basins coin-
cide with zones of coastal upwelling, the productive equato-
The logistic equation and its sigmoidal growth curve lead rial belt marks the gentle upwelling generated by the oppos-
to another important topic in biotic carbonate production ing effects of the Coriolis force in the northern and southern
– the different life strategies of organisms. As a rule, en- hemisphere, and the belt around Antarctica is caused by up-
ergy and other resources available to an organism are lim- welling within the Antarctic Current. Finally, the productiv-
ited and need to be partitioned between growth and repro- ity minima lie in the subtropical gyres where old, nutrient-
duction. In other words: organisms need to decide to either depleted, saline surface water is swept together by wind and