
Page 172 - The Biochemistry of Inorganic Polyphosphates
P. 172
WU095/Kulaev
WU095-08
Peculiarities of polyphosphate metabolism
156 March 9, 2004 20:32 Char Count= 0
(a) (b)
400 300
PolyP (mmol P (g dry biomass) −1 ) 200 PolyP (mmol P (g dry biomass) −1 ) 200
300
100
100
0
0 5 10 15 20 0 0 5 10 15 20
Time of growth (h) Time of growth (h)
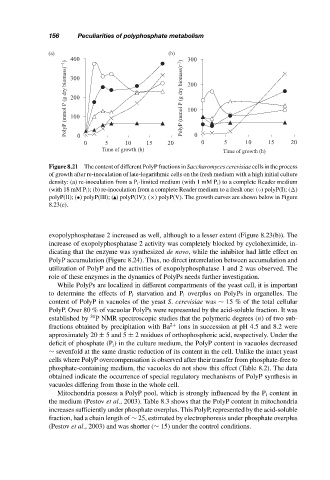
Figure 8.21 The content of different PolyP fractions in Saccharomyces cerevisiae cells in the process
of growth after re-inoculation of late-logarithmic cells on the fresh medium with a high initial culture
density: (a) re-inoculation from a P i -limited medium (with 1 mM P i ) to a complete Reader medium
(with 18 mM P i ); (b) re-inoculation from a complete Reader medium to a fresh one: (◦) polyP(I); ( )
polyP(II); (•) polyP(III); ( ) polyP(IV); (×) polyP(V). The growth curves are shown below in Figure
8.23(c).
exopolyphosphatase 2 increased as well, although to a lesser extent (Figure 8.23(b)). The
increase of exopolyphosphatase 2 activity was completely blocked by cycloheximide, in-
dicating that the enzyme was synthesized de novo, while the inhibitor had little effect on
PolyP accumulation (Figure 8.24). Thus, no direct interrelation between accumulation and
utilization of PolyP and the activities of exopolyphosphatase 1 and 2 was observed. The
role of these enzymes in the dynamics of PolyPs needs further investigation.
While PolyPs are localized in different compartments of the yeast cell, it is important
to determine the effects of P i starvation and P i overplus on PolyPs in organelles. The
content of PolyP in vacuoles of the yeast S. cerevisiae was ∼ 15 % of the total cellular
PolyP. Over 80 % of vacuolar PolyPs were represented by the acid-soluble fraction. It was
established by 31 P NMR spectroscopic studies that the polymeric degrees (n) of two sub-
fractions obtained by precipitation with Ba 2+ ions in succession at pH 4.5 and 8.2 were
approximately 20 ± 5 and 5 ± 2 residues of orthophosphoric acid, respectively. Under the
deficit of phosphate (P i ) in the culture medium, the PolyP content in vacuoles decreased
∼ sevenfold at the same drastic reduction of its content in the cell. Unlike the intact yeast
cells where PolyP overcompensation is observed after their transfer from phosphate-free to
phosphate-containing medium, the vacuoles do not show this effect (Table 8.2). The data
obtained indicate the occurrence of special regulatory mechanisms of PolyP synthesis in
vacuoles differing from those in the whole cell.
Mitochondria possess a PolyP pool, which is strongly influenced by the P i content in
the medium (Pestov et al., 2003). Table 8.3 shows that the PolyP content in mitochondria
increases sufficiently under phosphate overplus. This PolyP, represented by the acid-soluble
fraction, had a chain length of ∼ 25, estimated by electrophoresis under phosphate overplus
(Pestov et al., 2003) and was shorter (∼ 15) under the control conditions.